植物分子生理学グループの研究内容です
TEL. 086-434-1221
〒076-0046 倉敷市中央2-20-1
研究内容 (Research)SERVICE&PRODUCTS
佐々木孝行の研究テーマ
「植物におけるアルミニウム毒性と耐性機構の解析」
特に アルミニウム耐性遺伝子:ALMT輸送体,およびALMTファミリーの解析
根からの有機酸の放出は,Alを無毒化します.そこで、私たちはコムギから「Alによって活性化されるリンゴ酸トランスポーター(輸送体)」の遺伝子を単離し,Al耐性であり酸性土壌耐性の遺伝子であることを証明しました.
佐々木の研究は,このトランスポーターがどのように機能するか?発現がどのように調節されるかを調べることです.
次に,研究の背景とこれまでの研究の内容を紹介します.
詳しい内容 1「酸性土壌は作物の生育を阻害する典型的な"問題土壌"です!」
研究紹介にも書きましたが、この佐々木の研究で行ってきた主なテーマは,酸性土壌で問題になるアルミニウム(Alと略します)がいかに植物に影響するか?ということです.
では,酸性土壌って何なの?ということをご説明します.
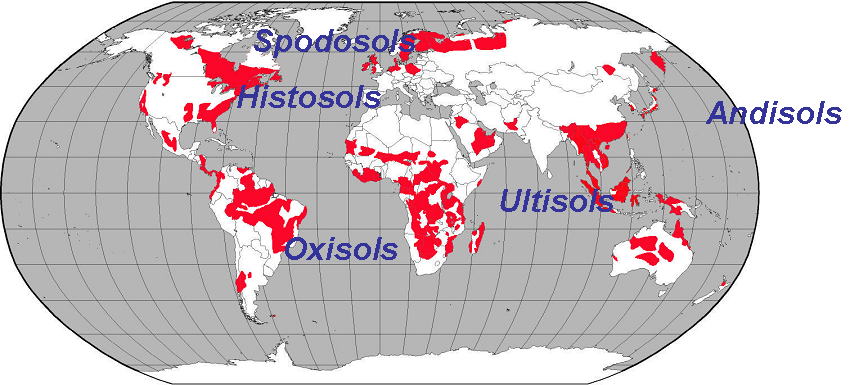
世界の農耕可能地の30~40%が,地図の赤で示す酸性土壌の地域となっています.
それぞれの地域で,異なる組成の酸性土壌であり,書かれているような名称で区別されています.
日本では,黒ぼく土(Andiisol)と呼ばれる火山性の酸性土壌が広く分布しています。

畑の土(中性)と黒ぼく土(酸性)(左の写真)で,エンドウを育ててみました.
すると播種後10日で,生育に差がでているのが分かります(右の写真)


エンドウを土から掘り起こしてみると,根の成長に違いがあるのが一目りょう然です.
この主な原因は,酸性土壌で溶け出してくるアルミニウムイオン(Al3+)が根の先端(根端)領域に集積するためです.
右の写真のようにAlが集積すると,根の伸長が阻害され,壊死がおこります.そして根が養分を吸収できなくなるため,植物の成長が抑制されてしまうのです.
詳しい内容 2「アルミニウムによる障害メカニズムを明らかにする!」
アルミニウムによる根の伸長阻害は,さまざまな要因によって引き起こされると考えられます.
そのうちの一つとして挙げられるのがAlによる活性酸素の誘発です.
タバコ培養細胞を用いて解析した結果,Alによる活性酸素の誘発がミトコンドリアの障害ならびにATPの減少とともに見られ,Alによる細胞死の原因の一つと考えられました(Yamamoto
et al. Plant Physiol. 128, 63-72, 2002).
このページでは,詳細は割愛します.
詳しく知りたい方は,植物成長制御グループのHPをご覧ください.
詳しい内容 3「アルミニウム耐性遺伝子を研究する!」
私たちは,コムギのアルミニウム耐性に着目して,Al耐性遺伝子の探索を始めました.研究に用いたコムギは,根端からリンゴ酸を放出することでAl耐性を獲得しています.
また,品種間でリンゴ酸の放出量が異なり,それによりAl耐性度が異なることも明らかになっていました.
私たちはAl耐性遺伝子を発見するために,準同質遺伝子コムギ系統のET8とES8を使用しました(下写真・左).
これらは,Al耐性系統(Carazinho)を供与親とし,感受性系統(Egret)を反復親として,8回の戻し交雑を行って作出されたものです(Delhaize et al. Plant Physiol. 1993, 103 685-693).
このように簡単に説明してますが,8回交雑を行うということは単純に8年間かけてつくりだされたコムギ系統ということです.彼らの努力には感服いたします...
そしてこのET8, ES8なくしてALMT1遺伝子の発見はなかった!と言っても過言ではないでしょう.
 左がES8,右がET8の圃場での生育です.
左がES8,右がET8の圃場での生育です.遺伝的背景が同じなので見た目にはほとんど区別がつきません.
さて,私たちはこのET8とES8の間で,根で発現している遺伝子を比較しました.
このときには,サブトラクション法という方法で,目的の遺伝子の探索を行いました.
(今だったら次世代シークエンスによるRNA-seq解析を行っていたことでしょう.)
その結果,ET8の根端で特異的に高く発現している遺伝子の単離に成功しました(Sasaki et al. Plant J. 2004, 37, 645-653).
これが「元祖」ALMT1(Aluminum-activated malate transporter 1)です.
現在は,他の植物のALMTタンパク質と区別するために,コムギの学名Triticum aestivumの略称と付けて「TaALMT1」とも呼ばれています(個人的にはThe ALMT1と書き続けたいのですが...).
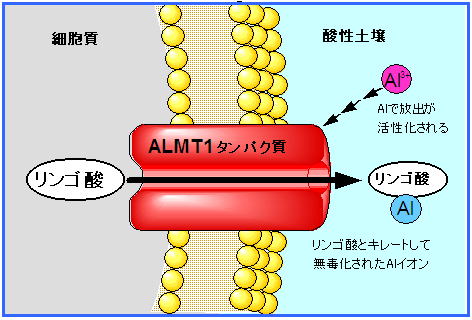
この論文(Sasaki et al. 2004)では,アフリカツメガエル卵母細胞を用いた二電極膜電位固定法や,形質転換植物など,さまざまな手法を用いて,ALMT1がAlで活性化されるリンゴ酸トランスポーター(注1)であることを証明しました.
そして,タバコ培養細胞を用いた系により,これがAl耐性に関わる遺伝子であることも証明しました.
(注1)現在,ALMT1はチャネル "channel"と考えられています.チャネルとトランスポーターは,イオンなどの輸送の機能に違いがあります.ではなぜ,トランスポーター "transporter" と呼ぶことになったのか. それは,「いろいろな事情」があったからです.
ALMT1遺伝子は,Al耐性系統の根端において常に発現しています.つまりALMT1タンパク質は常に根端に在るのです.
しかし,中性付近の土壌で,Al3+になっていないときにはリンゴ酸の輸送は活性化されません.
酸性土壌でAl3+が溶け出してくると,はじめて活性化されてリンゴ酸を根圏に放出するのです.
コムギは,なんとも効率的なAl耐性メカニズムをもっているものだ,と関心してしまいました。
ALMT1の発見以前にも,Alに応答して耐性に関与する(のではないか)という遺伝子は,いくつか報告されていました.
私たちの研究,リンゴ酸放出という生理的にAl耐性との関わりが解っている,その原因遺伝子の発見というのは「Al耐性メカニズムを遺伝子レベルで解明した」という意味でも,初めてであったと思います.

現在は,もう一つのAl耐性遺伝子として,クエン酸の輸送体がオオムギやソルガムなどから見つかっています。
これは,MATE (multidrug and toxic compound extrusion) と呼ばれるタイプの輸送体ファミリーが,Alで活性化されるクエン酸輸送を行うことが報告されています.
また,イネは非常に酸性土壌に強いのですが,コムギやオオムギらとはまた異なるAl耐性機構があることも解っています.
これらもまた,Al耐性遺伝子として近年注目されているものだと思います.
これらは,他の研究者の方がやっているので,それらの論文・総説をお読みください.
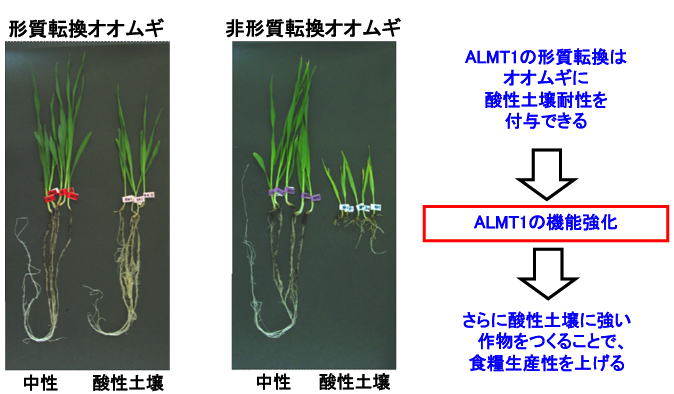
私たちは,ALMT1の発見を報告した同年に,ALMT1遺伝子をAl感受性のオオムギに発現させることで,Al耐性・酸性土壌耐性を付与できることも報告しました(Delhaize et al. PNAS 2004, 42, 15249-15254. ).
他にも,Al感受性のオオムギやコムギに発現させて,作物生産性が上がったという論文も発表されています(Delhaize et al. Plant Biotech. 2009, 7,391-400. Pereira et al. Annal. Bot. 2010, 106, 205-214.)).
すなわち,植物がもつ遺伝子によって酸性土壌耐性作物をつくりだせる可能性がでてきたわけです.
さて,コムギのALMT1遺伝子発現の量がAl耐性に重要なことは解っていただけましたでしょうか?
では,このALMT1遺伝子の発現量はコムギの根端でどのように制御されているのでしょう?
そこで私たちは,Al耐性度の異なる品種間で,ALMT1上流配列を比較してみました.
遺伝子の上流には「プロモーター」と呼ばれる,遺伝子発現を制御する領域が存在します.
私たちの解析により,コムギのTaALMT1遺伝子の上流には重複する配列領域があり,その重複数がAl耐性を制御する,ということでした(Sasaki et al. 2006).下の図はそのイメージです.
関連する論文もオーストラリアの共同研究者と論文になってます (Raman et al. 2008, Ryan et al. 2010).

そのほかにも私たちは,細胞膜局在や膜配向性(Yamaguchi et al. 2005, Motoda et al. 2007),コムギの主要なAl耐性遺伝子であり,4DL染色体に座上すること(Raman et al. 2005)を明らかにしてます.
ちなみに,コムギ(ここではパンコムギのことを言います)は,6倍体で A, B, Dゲノムが7つずつあります.
4DLというのは,下の図のように,Dゲノムの4番目,長腕(Long arm)にある,という事を意味します.

また,Al活性化機構やリンゴ酸を含むアニオン輸送体としての機能特性を電気生理学的に研究を行ってきました(Furuichi et al. 2010, Sasaki et al. 2014, Zhang et al. 2008).
Furuichi et al. (2010)では,アフリカツメガエル卵母細胞を用いた電気生理学的解析によって,TaALMT1タンパク質の C-末端にある比較的長い親水性ペプチド領域の3箇所の酸性アミノ酸{アスパラギン酸とグルタミン酸(E274, D275, E284)}がAl活性化に必要であることを明らかにしました.さらに,TaALMT1のC末端領域の欠失や,シロイヌナズナの相同タンパク質 AtALMT1のC末端領域と入れ換えたキメラタンパク質の解析から,C末端領域がリンゴ酸輸送能の調節に重要であることを明らかにしました.
Sasaki et al. (2014)では,同様に電気生理学的手法を用いて,ALMT1タンパク質のへリックス構造に着目し,N末端とC末端それぞれに,Al活性化に重要な機能ドメインが存在することを明らかにしました.
これらの結 果は,ALMT1の機能を強化する応用研究につながり,将来的にさらに強い酸性土壌耐性を作物に付与できると考えています.
ALMTの研究結果については,またアップしたいと思います。。。
注)文章中で、論文の雑誌名・巻・ページとかが省略されたものについては「研究業績・Publication」をごらんください。
詳しい内容 4「ALMTファミリー遺伝子の役割は? その1 気孔の開閉を制御する」
シロイヌナズナは,植物で一番初めに全ゲノム配列が明らかになったモデル植物です.シロイヌナズナには14(1つは配列が短くおそらく機能してないと推定されるので実質は13)のALMT相同遺伝子が存在します.そのうちAtALMT1もAlで活性化されるリンゴ酸トランスポーターであり,Al耐性遺伝子であることが分かりました (Hoekenga et al. PNAS 2006, 103, 9738-9743).
また,AtALMT9は液胞膜に局在するALMT相同遺伝子として報告されています(Kovermann et al. Plant J .2007 52, 1169-1180).AtALMT6も気孔の液胞膜にあるといわれています(Meyer et al. Plant J. 2011, 67, 247-257).
ただし,これら液胞膜タイプのALMTの生理的メカニズムは明確には示されていませんでした.
2013年に,AtALMT9が気孔の液胞に存在し,塩素イオンを液胞に取り込むことで,気孔開口に関与すると報告されました(De Angeli et al. Nature Commun. 2013,4, 1804).AtALMT9についての研究はスイスのグループがほかにも論文を精力的に出しています(Zhang et al. Plant Pysiol. 2013, 163, 830-843; Zhang et al. J. Biol. Chem. 2014, 289, 25581-25589).
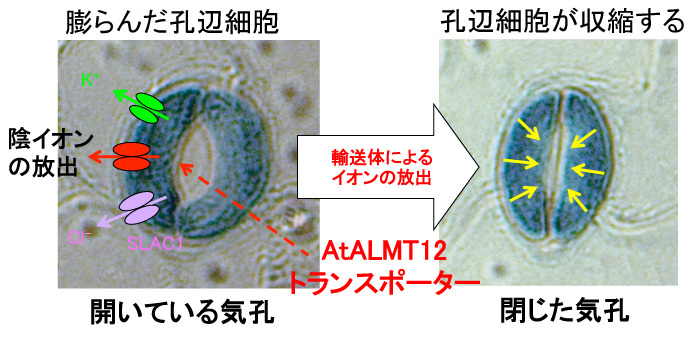
一方で私たちは,AtALMT12が気孔で特異的に発現し,気孔閉口を制御するアニオン輸送体の一つであることを報告しています(Sasaki, Mori, Furuichi et al. Plant Cell Physiol. 2010, Mar;51(3), 354–365).
気孔の閉口には、K+チャネルや陰イオンチャネル(SLAC1などが明らかになってます)が関与することが,以前から知られていました(詳しくは、同研究所・環境応答機構研究グループの森泉先生のHPなどを参考にしてください).
私たちのAtALMT12の論文は,気孔の生理機能にはこれらの輸送体以外にも「ALMTファミリー遺伝子」が関与することを明らかにした初めての論文です.
その後,AtALMT12が気孔のquick-type anion channelであると報告されました(Meyer et al. Plant J. 2010, Sep; 63(6) ,1054-1062).そのため,AtALMT12はQUAC1とも呼ばれています.
の研究には進展が報告されています(Mumm et al.2013、Imes et al. 2013).
私たちはAtALMT12による気孔閉口のメカニズムには,まだ不明な点が残されていると思ってます.
詳しい内容 5「ALMTファミリー遺伝子の役割 その2 果実の酸味に影響する」
果実の酸味は,リンゴ酸やクエン酸などの有機酸の量が関与します.
詳しい内容 6「ALMT研究の今後」
ALMTタイプ輸送体は,微生物や動物の既知のイオン輸送体とは相同性を示しません.つまり「植物特有の輸送体ファミリー」となっているのです.
近年のゲノム解析の進展により,シロイヌナズナ以外にも,イネやポプラなど多くの植物ゲノムデータベース上にALMT相同遺伝子が存在することが分かってきました.しかし,ほとんどの場合その輸送機能および生理的役割は不明です.
また面白いことに,当初は「Al活性化」される輸送体としてとられたALMTでしたが,これまでに報告されてきた多くのALMTタンパク質は「Al活性化」されないことも分かってきました.
このようなALMTファミリーの,Alで活性化されるもの・活性化されないもの,では何がその機能を決定しているのか?
このことも,今後明らかにしていきたいと思っています.
(Al活性化されないのを「ALMT」と言ってよいのか?とも考えています.ただ,発見された当時の機能に由来する遺伝子は数多く存在します.その意味では,一つの新たなトランスポーターファミリーとして「ALMT」の名前を残してもよいのではないかと思います)
ALMTが発見されて(正確には論文として報告されて)から,15年が経ちました.
今は紹介した他にも,いろいろな植物からたくさんのALMTが見つかり,それらの機能が明らかになってきてます.
植物生理学や植物栄養学,トランスポーター・チャネル研究の中で「ALMT」という名前が少しでも浸透してくれることを期待します.
バナースペース
植物分子生理学グループ
〒076-0046
倉敷市中央2-20-1
TEL 086-434-1221
FAX 086-434-1249