植物分子生理学グループの研究内容です
TEL. 086-434-1221
〒076-0046 倉敷市中央2-20-1
研究内容 (Research)SERVICE&PRODUCTS
佐々木孝行の研究テーマ
アルミニウム耐性遺伝子:ALMT輸送体,およびALMTファミリーの解析
次に,アルミニウム耐性遺伝子として発見したALMTファミリー遺伝子について行った研究内容をご紹介いたします.
詳しい内容 3「ALMTファミリー遺伝子の役割は?」
コムギのALMT1遺伝子を見つけたとき,生物の遺伝子データベースで,似たような遺伝子があるのかを検索しましたが,「unknown (知られていない)」遺伝子として登録されていました.つまり,それまでは遺伝子として何かのタンパク質をコードすることは解っていたのですが,何の遺伝子なのかが解っていなかったのです.さらに,ALMTの相同遺伝子は,植物にしか存在しませんでした.
詳しい内容 4「ALMTファミリー遺伝子の役割は? その1 気孔の開閉を制御する」
シロイヌナズナは,植物で一番初めに全ゲノム配列が明らかになったモデル植物です.シロイヌナズナには14(1つは配列が短くおそらく機能してないと推定されるので実質は13)のALMT相同遺伝子が存在します.そのうちAtALMT1もAlで活性化されるリンゴ酸トランスポーターであり,Al耐性遺伝子であることが分かりました (Hoekenga et al. PNAS 2006, 103, 9738-9743).
また,AtALMT9は液胞膜に局在するALMT相同遺伝子として報告されています(Kovermann et al. Plant J .2007 52, 1169-1180).AtALMT6も気孔の液胞膜にあるといわれています(Meyer et al. Plant J. 2011, 67, 247-257).
ただし,これら液胞膜タイプのALMTの生理的メカニズムは明確には示されていませんでした.
2013年に,AtALMT9が気孔の液胞に存在し,塩素イオンを液胞に取り込むことで,気孔開口に関与すると報告されました(De Angeli et al. Nature Commun. 2013,4, 1804).AtALMT9についての研究はスイスのグループがほかにも論文を精力的に出しています(Zhang et al. Plant Pysiol. 2013, 163, 830-843; Zhang et al. J. Biol. Chem. 2014, 289, 25581-25589).
一方で私たちは,AtALMT12が気孔で特異的に発現し,気孔閉口を制御するアニオン輸送体の一つであることを報告しています(Sasaki, Mori, Furuichi et al. Plant Cell Physiol. 2010, Mar;51(3), 354–365).
気孔の閉口には、K+チャネルや陰イオンチャネル(SLAC1などが明らかになってます)が関与することが,以前から知られていました(詳しくは、同研究所・環境応答機構研究グループの森泉先生のHPなどを参考にしてください).
私たちのAtALMT12の論文は,気孔の生理機能にはこれらの輸送体以外にも「ALMTファミリー遺伝子」が関与することを明らかにした初めての論文です.
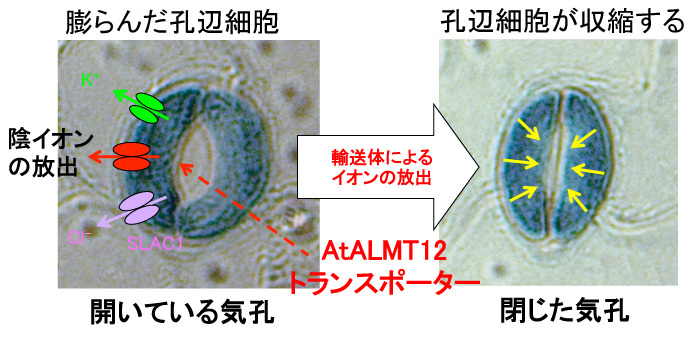
その後,AtALMT12が気孔のquick-type anion channelであると報告されました(Meyer et al. Plant J. 2010, Sep; 63(6) ,1054-1062).そのため,AtALMT12はQUAC1とも呼ばれています.
その後も,AtALMT12/QUAC1の研究で,進展が報告されています(Mumm et al. 2013、Imes et al. 2013).
しかい,私たちはAtALMT12による気孔閉口のメカニズムには,まだ不明な点が残されていると思ってます.
「2023年追記」
気孔閉口に関与するALMTは,シロイヌナズナで主に研究が進められてきました.
そこで私たちは,「他の植物でもALMT相同遺伝子が同様の機能を示すのか?」と疑問に思いました.
そこで,果実のモデル作物であるトマトをつかって研究を進めようと考えました.
(はなしが前後しますが,次の内容に書いているように,私たちはトマト(Micro-Tom)を使った研究を行っており,栽培のノウハウを知っていたという理由もあります)
そして,16あるトマトのALMT遺伝子の中から,孔辺細胞で発現するものがSlALMT11であることを明らかにしました.
AtALMT12は,アフリカツメガエル卵母細胞を用いた電気生理学的測定により,外液のリンゴ酸が存在するときにリンゴ酸放出が活性化されます(Meyer et al. Plant J. 2010, Mumm et al. 2013).
トマトのSlALMT11も外液のリンゴ酸で活性化される,リンゴ酸放出輸送体であることが解りました.
さらに,シロイヌナズナやトマトの比較によって気孔タイプのALMTの特徴として,C末端に酸性アミノ酸のグルタミン酸,アスパラギン酸が保存されていることが解ってきました.
このC末端酸性アミノ酸を欠損すると,気孔のALMTに特徴的な電流.電圧パターンが変化しました.
(通常は,マイナスを示す膜電位でリンゴ酸を放出しますが,一定の膜電位よりもさらにマイナス側になるとリンゴ酸の放出が抑制されます.しかし,酸性アミノ酸を欠損させると,よりマイナス側でのリンゴ酸放出の抑制がなくなり,リンゴ酸を放出し続けるようになりました)
この結果から,気孔タイプのALMTのC末端酸性アミノ酸は,膜電位センサーとしてはたらくと考えられました.
詳細は,Sasaki et al. Plant Cell Environ. (2022)で報告しています.
気孔の運動(開いたり,閉じたりすること))は,もう一つの陰イオンチャネル,カリウムチャネル,さらにプロトンポンプなどにより制御されています.気孔タイプALMTによる気孔運動の制御は,比較的最近になり明らかになってきました(Sasaki et al. 2010 が最初の報告です).そのため,気孔タイプALMTは他とは違う環境応答によって制御されている可能性があります.
そこで現在,気孔タイプALMTの特性について,さらに研究を進めています.
詳しい内容 5「ALMTファミリー遺伝子の役割 その2 果実の酸味に影響する」
果実の酸味は,リンゴ酸やクエン酸などの有機酸の量が関与します.リンゴ酸 (malate) は林檎(リンゴ;科名はMalus) に多く含まれるため,「リンゴ酸」と呼ばれるようになりました.
そこで私たちは,果実の酸味であるリンゴ酸の蓄積にも,ALMT輸送体が関与するのではないか,と考えました.
実験材料として,トマトを用いることにしました.
トマトは2012年にゲノム配列が解読され,果実のモデル植物ともいわれています.
(食品としてあつかわれる場合,日本ではトマトは果物ではなく野菜なのですが,植物学的には「果実」なのだそうです)
私たちは,トマトの中でそして丈が小さく大きくならない(セルフ・トッピング:self-toppingと言うそうです),Micro-Tom品種を用いることにしました.
日本でも,Micro-Tomを用いていろいろな研究をしているラボは多いと思います.
このMicro-Tomは,アグロバクテリウムによる遺伝子組み換えが可能で,サイズも小さいので植物インキュベーターなどの蛍光灯やLEDの光の下でも栽培が可能です.
私たちが,トマトからALMTを単離しようと試み始めたのが2010年で,当時はゲノム配列情報が十分ではありませんでした.
そこで,断片的に報告されているEST "Expressed Sequence Tags"という配列情報を元に,いくつかのALMT遺伝子をクローニングしました.
その結果,2つのトマト(Solanum lycopersicum)のALMT遺伝子(SlALMT)が果実で発現することを発見しました.
そして2012年にゲノム配列が報告され,トマトには「16」のSlALMT遺伝子の存在が明らかとなり,私たちがみつけていた果実のSlALMT遺伝子をSlALMT4とSlALMT5と名付けました.
(遺伝子の番号については,植物の場合,特に取り決めはなく,発見した順番だったり,染色体1番から座乗する位置の順番だったり,タンパク質のグループの中での順番だったり,様々です.トマトのSlALMTでは,私が最初に報告したため,染色体の位置の順番にしました)
SlALMT4とSlALMT5がコードするタンパク質は,クレード2とよばれるグループに分類されます.
クレード2には,液胞膜局在として知られているシロイヌナズナのAtALMT9が含まれます(Kovermann et al. 2007 Plant J.).
私たちは,この2つのSlALMT遺伝子に着目して研究を進めた結果,SlALMT4とSlALMT5はともに,果実において,種子につながる維管束で発現しており,さらにSlALMT5は発達中および完熟種子の胚で発現することを見いだしました.
 SlALMT5プロモーターにGUSを融合して発現させたトマト果実のGUS染色
SlALMT5プロモーターにGUSを融合して発現させたトマト果実のGUS染色青く染色されているのがSlALMT5の発現する維管束.萼(がく)の茎部分から種子へとつながるのが解る.
SlALMTとGFP融合したタンパク質を植物細胞(タバコ培養細胞とシロイヌナズナ葉肉細胞)のプロトプラストに一過的に発現させた結果,SlALMT4は小胞体(ER)膜に,SlALMT5はER膜とその他の細胞内膜系に発現することが解りました.
さらにアフリカツメガエル卵母細胞を用いた電気生理学的解析から,2つのSlALMTがリンゴ酸を透過することも解りました.
そして,SlALMT5については,遺伝子過剰発現トマトを作製しました.この遺伝子過剰発現系統と野生系統のトマト果実のリンゴ酸含量を比較しましたが,残念ながら差は見られませんでした.しかし,種子中のリンゴ酸(およびクエン酸)含量が有意に上昇することを見いだしました.SlALMT5は,種子につながる維管束や種子の胚で遺伝子発現がみられたことから,種子に有機酸を蓄積する働きがあると考えています.
(以上の成果は,Sasaki et al. Plant Cell Physiol. 57, 2367-2379 (2016) に掲載されています)
ちなみに,トマトを料理する際にトマトの「種(たね)」を取ることがあるようですが,それは種(とまわりのゼリー部分)に酸味があるからなのだとか.
おそらく,リンゴ酸やクエン酸が種に蓄積されていて,発芽時のエネルギーになるためではないでしょうか?
(食物の栄養学的な観点でトマトのことをあまり調べたことがないので,真偽のほどは今度調べます.すみません...)
さて,このSlALMT4, SlALMT5以外にも,トマトには残り14のALMT遺伝子が存在します.
そのうちの一つ,SlALMT9が液胞膜に局在し,トマト品種間でみられるリンゴ酸含量の違いに関与するという論文が2017年に報告されました(Ye et al. Plant Cell 2017 29, 2249-2268).
どうやら,SlALMT9の約400bp遺伝子上流の3塩基の挿入と欠損(indel)が遺伝子発現を制御することで,違いがみられるのだとか.
しかし,このSlALMT9遺伝子は,Micro-Tomや,ゲノム解読に用いられたHeinz 1706品種において,果実では発現がみられません(Sasaki et al. 2016より).でも,(成熟過程で変化しますが)リンゴ酸は果実に蓄積しています.
そのため,他のALMTや他の遺伝子も,果実のリンゴ酸含量に影響していると考えられます.
現在,まだ解明されていないトマトのSlALMT遺伝子にも注目しており,それらが生理学的にどのような働きをするのかについても調べていきたいと思っています.
詳しい内容 6「ALMT研究の今後」
ALMTタイプ輸送体は,微生物や動物の既知のイオン輸送体とは相同性を示しません.つまり「植物特有の輸送体ファミリー」となっているのです.
近年のゲノム解析の進展により,シロイヌナズナ以外にも,イネやトマトなどの作物や,ポプラなどの樹木でも,多くの植物ゲノムデータベース上にALMT相同遺伝子の存在が解ってきました.
しかし,ほとんどのALMT遺伝子については,その輸送機能および生理的役割は不明です.
また面白いことに,当初は「Al活性化」される輸送体としてとられたALMTでしたが,これまでに報告されてきた多くのALMTタンパク質は「Al活性化」されないことも分かってきました.
このようなALMTファミリーの,Alで活性化されるもの・活性化されないものがありますが,何がその差を決定しているのか?ますます解らなくなってきています.
このことも,今後明らかにしていきたいと思っています.
(Al活性化されないのを「ALMT」と言ってよいのか?とも考えています.しかし,機能が異なっても,最初に発見された当時の機能に由来する遺伝子名は数多く存在します.
たとえば,植物のCLC (chloride channel)は硝酸イオンを透過するものが多いそうです.(Barbier-Brygoo et al. 2011 Annu. Rev. Plant Biol. 62, 25-51)
このレビューでも紹介されていますが,「ALMT」ファミリーも,新たな一つのトランスポーターファミリーとして呼ばれているので,このままでも良いのではないかと思っています.
ALMTが発見されて(正確には論文として報告されて)から,15年以上経ちました.
今は紹介した他にも,いろいろな植物からたくさんのALMTが見つかり,それらの機能が明らかになってきてます.
植物生理学や植物栄養学,トランスポーター・チャネル研究の中で「ALMT」という名前が少しでも広まってくれることを期待します.
前のページに戻る


バナースペース
植物分子生理学グループ
〒076-0046
倉敷市中央2-20-1
TEL 086-434-1221
FAX 086-434-1249