
3.1. 葉肉コンダクタンス / Mesophyll condactance
二酸化炭素は気孔から葉内に入る
早朝,太陽が登る前,辺りは暗く植物はまだ光合成をはじめていません.日が出,光が差し始めると植物は光合成をはじめます.このとき光合成に必要な二酸化炭素が葉の中へと取り込まれます.葉の表面を覆うクチクラは二酸化炭素をほとんど透過しません.二酸化炭素は主に,開いた気孔を通じて取り込まれます.気孔から取り込まれた二酸化炭素はそのあとどのようにして光合成における炭酸固定の場である葉緑体のストロマに到達するのでしょうか.
気孔腔 / Substomatal cavity
一対の孔辺細胞とその周りを取り囲む副細胞で構成される気孔のすぐ内側には,気孔腔と呼ばれる空隙があります.この空隙は,液体で満たされているわけではなく,空気が存在しています.気孔腔の中の空気の湿度(水蒸気圧)は非常に高いとされており,研究者は100%の湿度になっていると考えています.一方,葉の外の湿度(水蒸気圧)は,気象によって異なります.晴れの日は湿度が低く,葉の中から気孔を通って水蒸気が葉の外へと出てきます.
ここでいう湿度は,相対湿度を指します.相対湿度は,温度によって変化します.また,葉の外の温度(気温)と葉の温度(葉温)は異なりますので,相対湿度を踏まえて水蒸気の移動を考えるのは,厳密に言うと正しくありません.一般的にはイメージしにくいかもしれませんが,空気中での水蒸気の運動は,湿度云々ではなく,水蒸気圧の高い方(水分子がより密集している方)から水蒸気圧の低い方(水分子がよりスカスカの方)へと移動すると考えるのが適当です.
気孔コンダクダンス / Stomatal conductance
光が当たることによって植物が光合成を開始すると,葉の中の二酸化炭素濃度が次第に低下するのにつれて,気孔腔の二酸化炭素の濃度が低くなってきます.すると,葉の中から水蒸気が出ていくのと同時に,二酸化炭素が葉の外から気孔を通って葉の中へと入ってきます.この時,二酸化炭素の取り込みの速度を決定するのは,気孔腔と大気の二酸化炭素の濃度差(二酸化炭素分圧)と気孔開度(より正確にはCO2に対する気孔コンダクタンス)です.空気中での二酸化炭素分子の運動は,二酸化炭素分圧の高い方から低い方へと移動します.気孔が完全に閉じていると気孔コンダクタンスが,最小(ほぼ0)になり,二酸化炭素は葉内に入りません.気孔が大きく開いていると気孔コンダクタンスが大きくなり,大気と気孔腔の間の二酸化炭素の分圧差により駆動されて,気孔を通じて葉内に入ってきます.
このような濃度に依存した物質の移動は,フィックの第一法則により記述することができます.
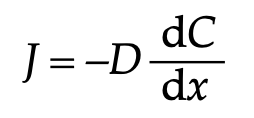
ここでJは二酸化炭素の移動速度です.
Dは二酸化炭素の拡散係数です.ここでは,Dは気孔コンダクタンスに相当します
xは距離を意味しています.気孔の穴の中に,大気から気孔腔までの二酸化炭素の勾配があると考えられます.ですから,ここでdxは大気から気孔腔に向かって,だんだん二酸化炭素の濃度が変わっていくあいだの距離を示しています.
そして,Cは二酸化炭素のモル濃度です.これは距離xにつれてだんだん変わっていきます.dC/dxはこの濃度の変化の割合を示していることになります.
気孔の穴の厚さはほとんど変わらないと考えて良いですから,二酸化炭素の移動速度(J)は,大気と気孔腔の間の二酸化炭素濃度差と気孔コンダクダンス(Dは一般的な拡散係数をしめす記号として使われています.気孔コンダクタンスは–Dの代わりに,gsと表記されることが多いです.gはコンダクタンス(伝導度)sはStomata(気孔)を意味しています)によって決まるということをしめす数理モデルを考えることができます.
気孔腔から葉肉細胞そしてストロマへ
気孔腔に到達した二酸化炭素は,二酸化炭素の化学ポテンシャル勾配に従って,葉肉細胞の細胞壁を取り囲む水の層の中に溶け込みます.水に溶け込んだ二酸化炭素は,さらに,細胞壁の繊維の間の水を通り抜けて進みます.そして細胞の外側を包み込む原形質膜へとたどり着きます.原形質膜は,脂質二重膜で構成されているため,疎水的な二酸化炭素分子は透過しやすいと考えることができますが,実際に測定をすると,どうも人工的な脂質二重膜とは異なり実際の原形質膜は二酸化炭素をあまり透過しないようです.これは,非常にタンパク質が多いということが原因かもしれません.二酸化炭素分子は,なんとかかんとか葉肉細胞の原形質膜を通過すると細胞質基質にたどり着きます.
細胞質基質から,葉緑体に入っていくに際して,原形質膜から葉緑体までの距離は大変大きな問題です.二酸化炭素は,空気中ではその拡散速度が早いですが,水中では極端に遅くなります.ですから,できるだけ水の層を通り抜ける距離は短い方が,素早く辿り着けるのです.その理屈ですと,原形質膜から葉緑体までの距離が近ければ,水を含む層である細胞質基質のなかを二酸化炭素が拡散する時間が短くなり,素早く葉緑体に二酸化炭素が供給されることになります.
葉緑体に接近した二酸化炭素分子は,葉緑体の包膜(外包膜と内包膜)を通過し,とうとう葉肉細胞内の葉緑体の中のストロマに到達します.このシナリオは,気孔腔に隣接する葉肉細胞に焦点を当てたものです.気孔腔から離れた場所にある葉肉細胞では,細胞壁の中,あるいは他の葉肉細胞を通り抜けて二酸化炭素が到達するため,このような移動をなんども繰り返していく必要があります.細胞壁の厚さ,原形質膜から葉緑体までの距離,原形質膜の二酸化炭素のとおり易さなど,気孔腔からストロマまで二酸化炭素が移動していくまでの通り易さにはいろいろな要因があります.気孔腔からストロマまでの二酸化炭素の透過し易さは葉肉コンダクタンスと呼ばれます.
気孔開度でほぼ一義的に決定される気孔コンダクタンスと異なり,いろいろな要因が関連する葉肉コンダクタンスですが,気孔腔からストロマまでの二酸化炭素の移動も,濃度に依存しています.このため,気孔コンダクタンスもフィックの第一法則により記述することができます.
気孔コンダクタンスと葉肉コンダクタンスで異なる点はdC/dxが,大気と気孔腔の二酸化炭素の濃度勾配ではなく,気孔腔とストロマの二酸化炭素濃度勾配に相当するという点で,基本的には同じ数理モデルを考えることができます.そして,記号としては,gmと表記されることが多いです.mはmesophyll(葉肉細胞)を意味しています.もちろんこれまで解説してきたように,コンダクタンスを構成する要因はgsとgmではだいぶ異なります.
また,気孔コンダクタンスの場合はほぼ一方向に向かって二酸化炭素が移動するので,フィックの第一法則がそのままモデルとすることができますが,葉肉コンダクタンスは三次元的にまた不均一に二酸化炭素が拡散しますので,一次元のフィックの第一法則が数理モデルそのまま成り立つというわけではありませんが,気孔腔からストロマまでの二酸化炭素の拡散の詳細を把握するのは現実的ではありません.葉全体のストロマの二酸化炭素濃度の平均と考えられる値を求め,また葉全体の気孔腔の二酸化炭素濃度の平均と考えられる値を求め,そこから葉全体の葉肉コンダクタンスの平均と看做してよい値を計算により求めるのが実際の葉肉コンダクタンスの測定の原理となっています.
3.2. アクアポリンとCO2輸送 / Aquaporins and CO2 transport
アクアポリン
原形質膜の二酸化炭素透過性が実際には意外に低いと考えられていることを紹介しました.葉肉細胞の二酸化炭素透過性は,アクアポリンにより調節されているというモデルが提唱されています.光があたり光合成をするときなどは,アクアポリンの活性調節により,二酸化炭素透過性が上昇していると言う考え方です.その根拠となる研究がいくつかあります.しかし,それらの報告の中には争論となりそうな文献もあるため,ここではあえて触れないようにします.研究者の間では,葉肉コンダクタンスの調節に二酸化炭素を透過する能力のあるアクアポリンが関与しているという作業仮設のもと研究を進めているのが流れになっていますが,根拠をもって結論づけるのは時期尚早で,研究は途上であるといえます(2023年5月記).
COOポリン
一方,数あるアクアポリン分子の中に,水分子以外に二酸化炭素を透過するものがあるのは,確実です.私自身の論文でもこのことについて報告しています.論文査読の際に,二酸化炭素輸送活性測定の再現性が取れないとの批評をいただいたことがあります.測定方法の中で,低二酸化炭素溶液から高二酸化炭素溶液へとすばやく入れ替える手順が必要になりますが,ここが二酸化炭素輸送活性を測定する場合の技術的な大きなポイントのひとつです.実験がうまくいかない理由の全てがこのポイントであるとは言いませんが,実験の再現性において,ここは非常に決定的なポイントです.
いずれにせよ,アクアポリンの全てではなく,その一部が二酸化炭素を受動的に輸送することを示す実験証拠はしっかりとしています.アクアポリンの中で二酸化炭素を透過するものについて,COOポリンと呼称することが提案されています.一方,そのCOOポリンが,gm調節に関わっているか?という話題になると,そこは現在進行中の課題です.
私は,CO2を輸送するアクアポリンと輸送しないアクアポリンの構造を比較することで,アクアポリンのCO2輸送活性に必要なアミノ酸を同定する研究を進めています.これによりアクアポリンの構造とCO2輸送活性の関連が明らかとなり,効率的な光合成に必要な葉肉コンダクタンス調節をより詳細に理解することができます.
このテーマに関連する代表的な論文
- Izumi C. Mori, Jiye Rhee, Mineo Shibasaka, Shizuka Sasano, Toshiyuki Kaneko, Tomoaki Horie, Maki Katsuhara
CO2 transport by PIP2 aquaporins of barley
Plant and Cell Physiology 55: 251–257 (2014)
doi.org/10.1093/pcp/pcu003