
* 細胞壁形成関連変異 shv3
シロイヌナズナのトランスポゾン挿入変異ラインを利用して、細胞壁形成変異株shv3を分離しました。この変異株は元々根毛ができない表現型で選抜されたのですが、詳細な解析からこの変異株では細胞壁の形成に異常があることがわかりました。類似遺伝子SVL1の破壊変異の多重変異株では、根毛以外にも様々な表現型が見られました。原因遺伝子は植物特有の遺伝子で、今後遺伝子の機能が明らかにされれば、細胞壁形成について新たな知見が得られることが期待されます。
根毛の様子 shv3svl1二重変異株の胚軸の様子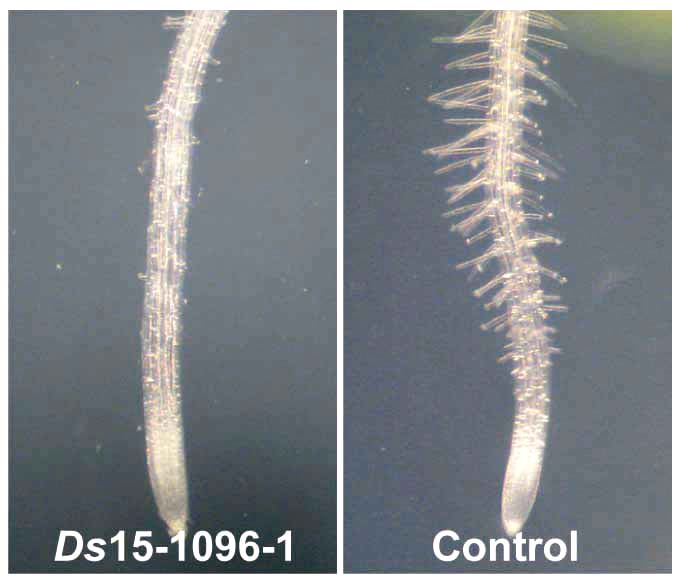
*クチクラ層形成関連変異 cof1
トランスポゾン挿入変異ラインから葉が癒着する変わった変異を見つけました。詳細な解析からこの変異株は、植物細胞の表皮を乾燥や病害虫からの攻撃から守 るクチクラ層の形成に関わる因子であることがわかりました。COF1は膜を介した物質の輸送に関わっているABCトランスポーターと呼ばれる一連のタンパ ク質のグループに属し、解析の結果から脂質の輸送に関わっていることが示唆されています。
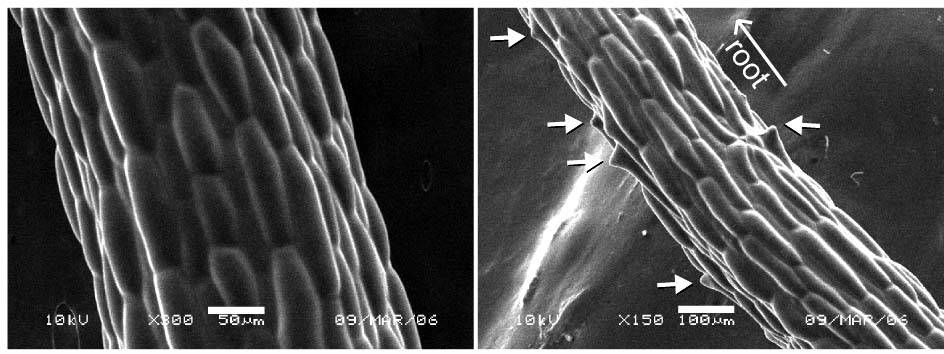
植物体の様子 走査電子顕微鏡による表皮の観察像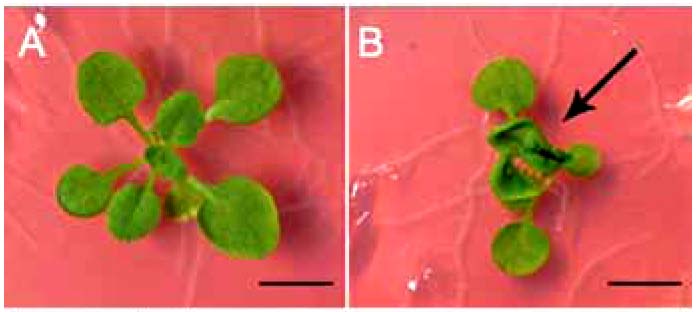

左図: 野生株(左)とcof1 変異株(右)の植物体の様子 (bar=10 mm)。cof1変異株では本葉が融合している。右図: 走査電子顕微鏡による表皮の状態 (bar=5 μm)。野生株(左)では結晶化したワックスの構造体が見られるが、cof1変異株 (右) ではほとんど見られない。
*エタノール高感受性変異 gek1
変異株を収集している過程でエタノールに高感受性を示す変異株を取得し、geko1 (下戸1) と名付けました。geko1 (gek1) は、野生株の100倍程度強くエタノールに感受性を示します。他のアルコールへの応答や、他のストレスへの応答には影響がありません。また、通常の生育にも何ら 影響は認められていません。解析の結果、エタノールから作られたアセトアルデヒドに感受性を示していることが明らかとなりました。geko1変異の原因遺伝子を同定したところ、植物と古細菌にのみ見いだされ る今までに報告のない遺伝子であることが判明しました。その後、GEK1はD-aminoacyl-tRNA-acylaseの活性を持っていることが報告されました。この活性とエタノール感受性との関係はまだ不明です。今後の解析により、これまでに知られていない植物が持つ秘密がわかるかもしれません。
エタノール存在下での発芽試験 GEK1高発現植物体 のエタノール耐性

左図:0.01%という低濃度でも、geko1変異株(gek1-1, 1-2, 1-3)では、発芽が著しく阻害される。 (Hirayama et al. PCP, 45, 703- )。右図:3% エタノール存在下で生育させた野生株(左)とGEK1高発現形質転換体植 物(右)(Fujishige et al. PCP, 45, 659-)。
>>> アブシジン酸応答機構の解析
>>> データ駆動型作物デザイン技術の開発