|
   

植物と善玉・悪玉ウイルスの相互作用を紐解く
植物の生育は様々な微生物との相互作用に大きな影響を受けています。植物に病気を引きおこすウイルス、細菌、糸状菌はその生育に脅威となります。一方、悪玉である植物の病原体に感染し、生物農薬として有効利用できそうな善玉ウイルスも見つかっています。本グループでは、ウイルスの多面性に着目し、二者(植物-ウイルス)、三者(植物-病原糸状菌-マイコウイルス)間の多様な相互作用・鬩ぎあいの研究を進めています。最終目的は、植物の健全な育成を図ることにあります。


ウイルス研究から、作物を病害・虫病害から守るあらたな仕組みを作り出す
准教授: 近藤 秀樹
E-mail:hkondoATokayama-u.ac.jp
Pure-Elsevier / researchmap
■農生態系におけるウイルス動態を理解する
作物の生育はそれを取り巻く微生物、昆虫類により様々な影響を受けています。作物自身やそれを取り巻く微生物、昆虫類には多様なウイルスが存在していますが、それらの作物生産への影響はまだ良く理解されていません。そこで、作物の生存圏におけるウイルスの多様性や感染動態を解析し、作物生産へのインパクトを理解することを目指しています。→得られた知見を基盤としたウイルス病害の制御やウイルス自身を利活用できると期待されます。
最近の主要成果
コムギに感染する新規ベータフレキシウイルスの発見. Front Microbiol 12, 715545, 2021
オオムギの重要害虫であるアブラムシのウイルス叢を解明. Front Microbiol 11:509, 2020
菌類ウイルスと植物ウイルスの間に見出された双方向の促進的な相互作用. PNAS 117:3779-3788, 2020
生物界(植物・菌類)をまたいだウイルスの自然界での発見! PNAS 12267-1227, 2017
以下をご参照ください.
植物・菌類ウイルスの生物界をまたぐ感染.Annu Rev Virol . 2023 10(1):119-138
菌類ウイルスは動物に感染できる? PLoS Pathog. 2023 19(10):e1011726
分節型ケノムを持つラフトウイルス . ウイルス 63 号143-154, 2013. J-stage へ
ネコブカビ類により媒介される土壌伝染性ウイルス JGPP 79, 307-320, 2013. link
テンサイそう根病の病原ウイルス(BNYVV)の進化と品種抵抗性. 植物防疫 68, 168-179, 2014

 研究内容をもっと詳しく 研究内容をもっと詳しく

病原体感染戦略の弱点を理解し、作物を病害から守る
准教授: 兵頭 究
E-mail:khyodoATokayama-u.ac.jp
Pure-Elsevier / researchmap
新学術領域「ネオウイルス学」 インタビュー記事
■植物ウイルスの複製の仕組みを紐解く
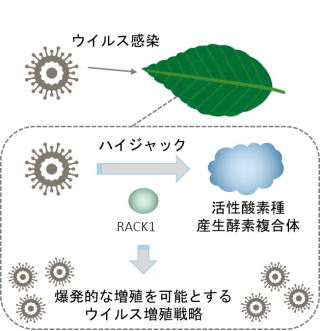
植物ウイルスの多くは複製を効率的に進めるため、宿主細胞内にウイルス合成工場を誘導します。そこには多くの宿主因子(蛋白質)も含まれています。ウイルスはそれらの因子を有効に利用し、効率的に複製することが明らかになりつつあります。→ウイルス複製の仕組みを理解して、ウイルス病の制御に利用することが期待されます。
最近の主要成果
植物ウイルスによる宿主ハイジャックの新戦略を解明.New Phytologist 221: 935?945, 2019
植物ウイルスの複製に宿主ROS産生系を転用する. PNAS 114,E1282-E1290, 2017
以下も参照ください.
植物RNAウイルスの複製・翻訳にかかわる宿主因子. Current Opinion in Virology 17, 11-18, 2016
coming soon
WTT助教:佐藤 有希代


名誉教授 : 鈴木 信弘
E-mail:nsuzukiATokayama-u.ac.jp
Pure-Elsevier / Loop-Frontiers/ Google scholar/ researchmap/ 岡山大リポジトリ
ヴァイロコントロール:ウイルスをもってカビを制す
作物にはいろんな病気にかかりますが、その被害により10億人分の食料が失われていると推定されています。中でも菌類による被害はその80%を占めると言われています。その菌類による病気をウイルスを使って防ごうというアプローチがヴァイロコントロールとなります。
ウイルスは生きとし生けるものすべてから見つかりますが、ほとんどのウイルスは病気を起こしません。菌類に感染するウイルスも然りで、一部ウイルスが宿主菌類に病気を起こします。この菌類を病気にする例外的なウイルスがヴァイロコントロールの主役になります。例えば、クリを殺してしまうクリ胴枯病菌に対してはハイポウイルスという救世主が現れ、ヨーロッパのクリ樹を守っています。現在、日本の果樹を恐ろしい白紋羽病菌(子のう菌)から守るため、救世主となり得るウイルスの探索、性格付けを進めているところです。最近、有望なウイルスも見つかり、実験室レベルでのウイルスの防除効果は実証されています。
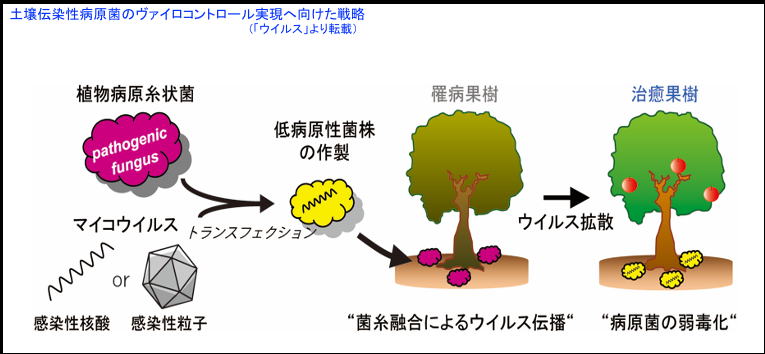
菌類ウイルスの分子生物学 (植物/病原糸状菌/ウイルス) :クリ/クリ胴枯病菌(子のう菌)/ウイルス、果樹/紋羽病菌/ウイルスの系を使い、菌類ウイルスの病徴発現、複製・伝搬機構の細胞レベル、分子レベルでの解明を進めています。これらの成果に基づき菌類ウイルスの生物防除因子としての増強・利用を図り、ウイルスを用いた植物病原糸状菌の防除法(ヴァイロコントロール)の実用化を目指しています。
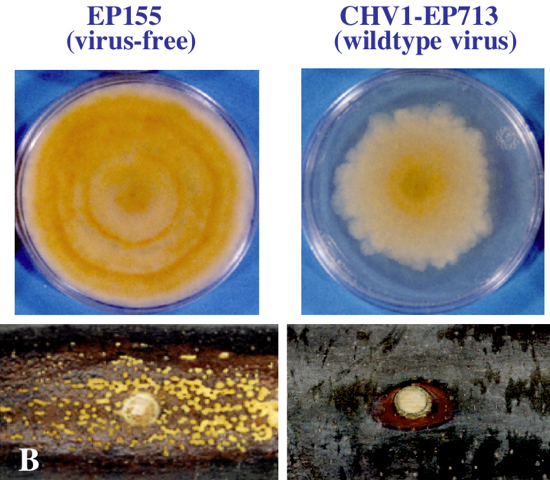 
ハイポウイルスに感染したクリ胴枯病菌とクリに対する病原性
ネオウイルス学の構築 これまでのウイルス学は植物あるいはヒトを含む動物に病気を起こすウイルスの研究から発展してきました。本研究室では、これまで見過ごされてきた植物が絡む生態系での菌類ウイルスの役割の解明、新たなウイルスライフスタイルの発見を目指しています。
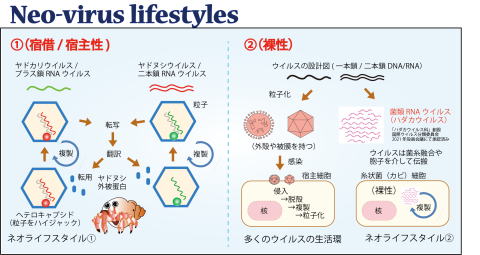
最近の主要成果
新しいウイルスライフスタイル(ヤドヌシ・ヤドカリ性). Nature Microbiology 1, 15001, 2016. link / リポジトリ1 / リポジトリ2 (Virus Res 2016)
Hadaka virus 1のユニークな特性(裸性)を報告しました.mBio 11, e00450-20, 2020
DICERは翻訳,転写れべるで抗ウイルスRNAiに関与する. PNAS 116 2274-2281, 2019
抗ウイルスRNAi機構のSAGA複合体により転写制御. PNAS 114, E3499-E3506, 2017
菌類ウイルスゲノムで初めてのIRES(リボゾーム内部進入部位)の報告. mBio 9, e02350-17, 2018
以下を総説もご参照ください.
マイコウイルスの多様性と進化について.Annu Rev Phytopathol. 2022 60:307-336
マイコウイルス研究の新展開 日本植物病理学会報 83, 129-132, 2017. link
マイコウイルスとヴァイロコントロール ウイルス,60, 163-176, 2010. J-Stageへ
マイコウイルス宿主としてのクリ胴枯病菌. ウイルス 64, 11-24, 2014 . J-stage へ
多様性に満ちた菌類2 本鎖RNAウイルス. ウイルス 64, 225-238 J-stage へ
研究内容をもっと詳しく
最近の研究助成 (科研費 2013〜)
2025〜2028 【開 拓】 ネオウイルス学から現代ウイルス学への挑戦
2025〜2028 【基盤B】 植物ウイルス感染における宿主オートファジー経路の正・負の役割の解明
2025〜2027 【萌 芽】 植物ウイルスの細胞侵入機構の理解に向けたウイルス検出系の開発
2023〜2026 【基盤B】 作物超個体における根圏RNAウイルス叢の実体解明とその生態学的役割
2021〜2025 【基盤S】 マイコイミュニティ研究の最前線とその植物病理学への新展開 【中間評価結果】
2023〜2025【萌 芽】 絶対寄生菌ウイルスの研究プラットフォーム構築と生物防除への挑戦
終了課題
2021〜2024 【開 拓】 植物・菌・動物の生物界を跨いだウイルス感染:宿主との攻防と適応戦略
2021〜2023 【基盤B】 MAPKカスケードを巡る植物−ウイルス間相互作用の解明
2022〜2024【萌 芽】 実験室進化から紐解く抗ウイルス免疫打破株の出現と耐性機構
2022〜2024【二国間交流事業】 クリ樹体内での多様なウイルス(感染クリ胴枯病菌)の挙動と生物防除能
2022〜2024【特別研究員奨励費】 3つの生物界を跨いだウイルス感染
2021〜2022 【萌 芽】 モデル糸状菌アカパンカビを用いたウイルス研究フロンティア研究 (挑戦的研究 (萌芽))
2021〜2021 【基盤A】 マイコイミュニティ研究の最前線とその植物病理学への新展開
2019〜2021 【基盤B】 作物根圏におけるウイルス叢の多様性とその感染動態から紐解 く生態的意義 (基盤B)
2019〜2020 【若手B】 ROSを介した植物ウイルスタンパク質の機能制御機構
2019〜2020 【新学術/研究領域提案型】 植物ウイルス:細菌/糸状菌感染における制御因子
2017〜2020 【基盤A】 植物病原糸状菌の抗ウイルス免疫機構研究のフロンティア
2016〜2020【新学術/研究領域】「ネオウイルス学」 糸状菌ウイルスのネオ・ライフスタイル
2015〜2017 【挑戦的萌芽】 ミトコンドリア感染性ウイルスの感染機構の解明と有効利用
2015〜2017 【基盤C】 メタゲノム解析に基づく菌類ウイルス叢の解明と有効利用に関する研究
2015〜2016【スタート支援】 植物免疫機構の転用によるウイルス誘導性免疫不全
2013〜2016【基盤A】 植物病原糸状菌の抗ウイルス自然免疫機構
2012〜2014【挑戦的萌芽】 アカパンカビモデル糸状菌を用いたウイルス/宿主相互作用解析系の確立
2012〜2014 【基盤C】 植物ゲノムに存在する非レトロウイルス様配列の挿入メカニズムと病理学的意義
2021〜2022【特別研究員奨励費】 クリ胴枯病ヴァイロコントロール因子の宿主範囲決定因子
2018〜2019【特別研究員奨励費】 ヤドカリウイルスの新奇生活スタイル(宿借性)の証明
2015〜2017【特別研究員奨励費】 新ウイルスライフスタイル:dsRNAウイルスを宿主とする裸のssRNAウイルス
2013〜2014【特別研究員奨励費】 白紋羽病菌のヴァイロコントロールに向けた新規2分節dsRNAの性状解析
2011〜2013【特別研究員奨励費】 ハイポウイルスの多機能性蛋白質p29により誘導されるレオウイルスゲノム再編成
主な民間研究助成
上原記念生命科学財団 平成27年度研究助成
八雲環境科学振興財団 平成27年度研究助成
両備てい園財団 第35回研究助成
山陽放送学術文化財団 平成24年度研究助成
研究室、研究設備の紹介

研究室は一号館二階の真ん中
 
廊下もすっきり/ウイルスチームのメイン実験室(きれいに、そして機能的になりました)。
-例えばこんな機器を利用しています-
透過型電子顕微鏡(TEM;共通機器)/ラボのPCRはいつもフル稼働!
 
超遠心分離機(共通機器) ぐるぐる回ってます/共焦点レーザー顕微鏡 (PMI所有)
 
育成室で実験植物がすくすく育っていま/温室(入り口側1/3をPMIが利用しています)
 
実験室からみた研究圃場(オオムギ)



|

